Evolution & Systematics
Through out history the phylogeny of Echinoderms have been controversial because of morphological characteristics being used, with multiple possible evolutionary pathways being found dependent on the characteristic (Smith, 1988; Raff et al., 1988). In trying to understand the evolutionary pathways from which Echinoderms have evolved the grouping and order in which the classes evolved is important. Raff et al. (1988) after molecular analysis developed the theory that ophiuroidea was the sister group to crinoidea (Fig.1). In Cohen et al. (2004) recognized that asteroidean are the closest sister group to crinoids (fig. 2). Dependent, up on which DNA sequence, morphological characteristics and are more highly favoured, defines the relatedness of the classes in this phylum.

Figure 1: Adapted from the Boolootian (1966). Early understanding of Echinoderm class phylogenetic relationship. In this phylogenetic tree shows the early divergence of the other class from crinoids.
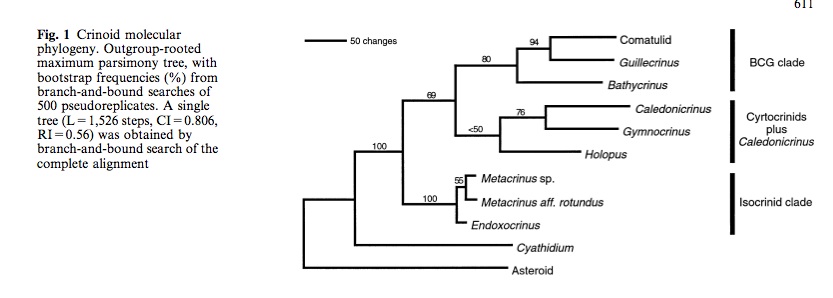 
Figure 2: Recent phylogentic tree showing the sister group of crinoidea as the Asteroids. The tree also displays the three clades within the class of crinoidea. Comanthus gisleni is found within the BCG clade. (Cohen et al.,2004)
Homologous characteristic have been refined using molecular sequence and 18s RNA subunit to estimate phylogenetic relatedness (Raff et al.,1988). This region of RNA is highly conserved acrossand within species making it useful in distinguishing relatedness between outgroups and classes(Raff et al.,1988). Theresults Raff et al. (1988) study yielded the class of Echinodermata evolved from a crinoid ancestral form (fig. 1).
The ancestral form of crinoids were sessile and stalked (Cohen et al.,2004, Boolootian,1966). They diverged from the echinoderm common ancestor at the end of the Lower Cambrian (550 Ma) (Raff et al.,1988). Evolution of extant morphological characteristics evolved from four genera in the upper part of the Lower Cambrian (fig.3). In the “Physical Description”section when the locality of the mouth and anus were discussed and shown, it can be linked to the evolution and divergence of Camptostoma genera (Fig. 3). Other characteristics found in extant genera can be linked in the fossil record including the calyx and tegmen on the central dorsal plate (Smith, 1988).
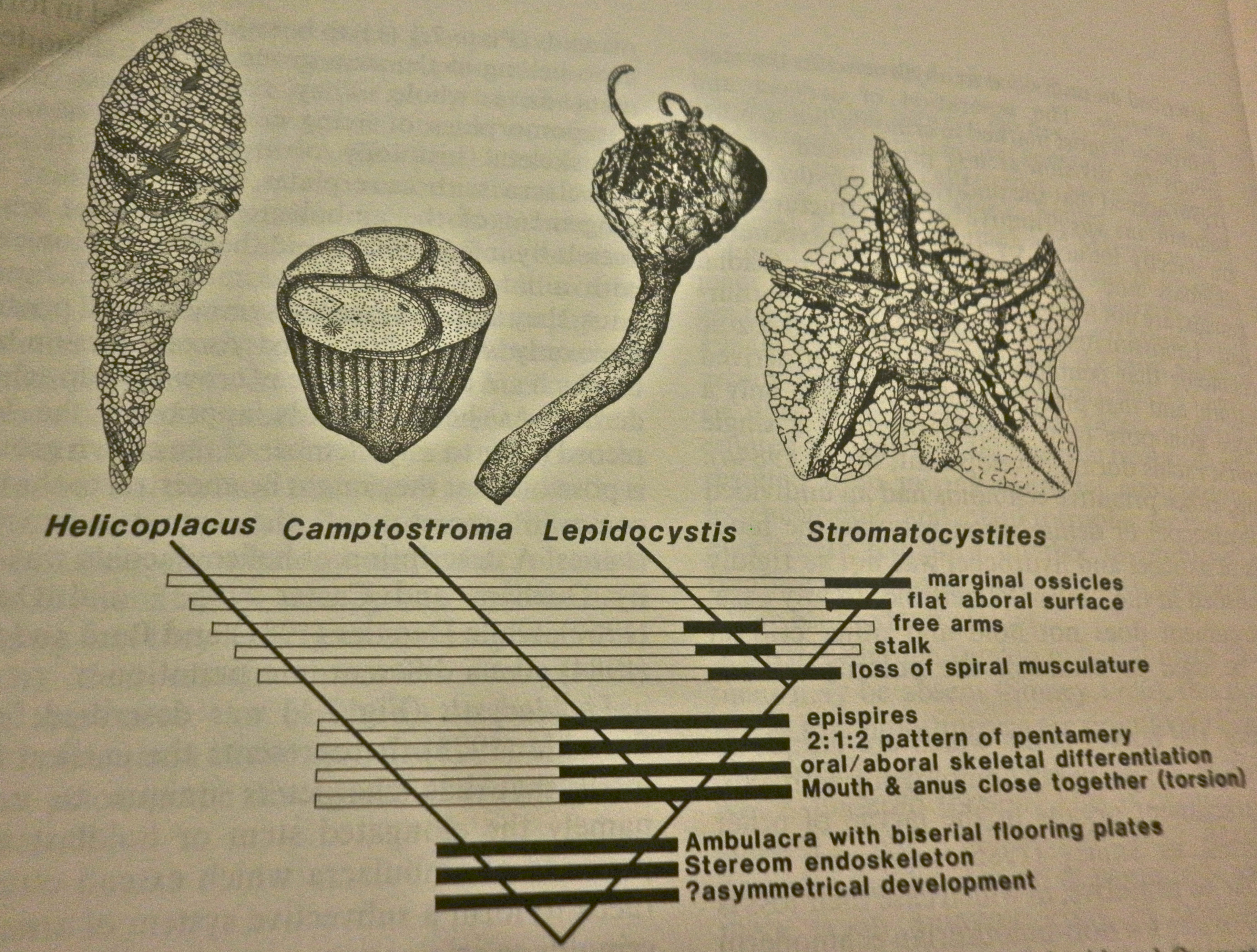
Figure 3: Four ancestral genera that have the extant morphological characteristics. (Smith, 1988)
Modern molecular analysis of the class Crinoidea has divulged three clades within this class (fig. 2). Comanthus gisleni is found within the BCG clade – Bathcyinus, Comatulid and Guillecrinus (Cohen et al.,2004). This clade includes the derived stalked crinoids, commonly referred to as “stalk-less”. However, is a common misconception because the central dorsalplate on which the ciri is attach, ontogenetically is the last remaining segment of the stalk (Cohen et al.,2004).
The stalk-less crinoids other common name is “feather-star”, this includes 140 genera and 500 species (Messing, 1997).
|